





以下文章來源于一麥眾承 ,作者倪永靜
作者簡介:
倪永靜,1983年,2010年畢業(yè)于河南農業(yè)大學,作物遺傳育種專業(yè)。2010年7月——至今,在商丘市農林科學院小麥研究所工作,從事小麥常規(guī)育種和遺傳育種。與河南農業(yè)大學合作選育的小麥新品種“尚農6”“尚農8”于2022年6月通過河南省主要農作物審定委員會審定。另外在試品種7個。
1創(chuàng)建小麥突變體庫
選用均勻一致的國麥301種子1kg,首先在冷水中浸泡4小時,然后用0.4%(v/v)EMS溶液在0℃處理種子2小時,接著在20℃處理14小時。排干EMS溶液,在流水中沖洗4個小時,晾干后種植于商丘市農林科學院雙八試驗基地。M?代種子于2012年10月單粒播種,行距0.23m,株距0.08m,按單穗收獲(即M?種子);M?種子于2013年10月種穗行,行距0.23m,株距0.08m。M?、M?均以未處理的國麥301為對照。田間管理按一般試驗田進行。
每7~10d田間調查一次,觀察記載幼苗、葉形、莖、分蘗、穗部、育性等性狀,篩選各種表型突變體。變異穗系全部單株收獲脫粒后,觀察記載籽粒形狀、大小、顏色等性狀。
對M? 12020個穗系的主要農藝性狀及其他生物學性狀進行調查,共獲得769個突變體系。其中,5個苗期致死、109個株高突變、177個葉片性狀突變(幼苗期、成株期)、82個穗部性狀突變、18個成熟期發(fā)生突變,64個分蘗突變,45個不育突變,52個籽粒突變,217個其他性狀的突變。M?中發(fā)生表型突變的頻率約為6.398%。具體突變類型如表1,部分突變體類型展示如圖1。
表1 EMS 誘變小麥新品種國麥301 M?田間突變類型

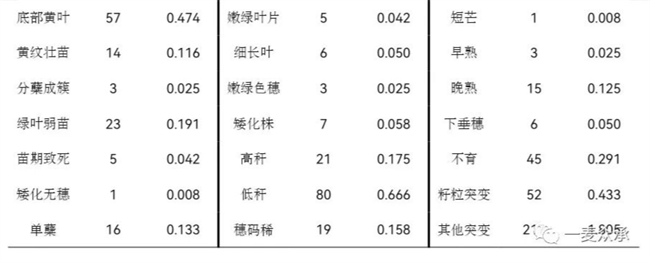

























圖1部分突變體類型
2 矮化獨稈突變體dmc的生物學特征及遺傳分析
?2.1獨稈突變體的獲得及其生物學特征
通過連續(xù)單株選擇,獲得了1個不分蘗突變體dmc純合株系。該突變體完全不分蘗,只有一個主莖(圖2-A),個別植株偶爾會有1個小分蘗。2016-2017年大田突變體株系dmc平均分蘗1.11個,突變親本國麥301平均分蘗21.73個。突變體dmc植株明顯矮小,株高48.00cm,是突變親本國麥301的74.53%(圖2-A)。進一步對比發(fā)現(xiàn),突變體dmc只有4個節(jié)間,比國麥301少了一個節(jié)間,與突變親本國麥301相比,突變體dmc各節(jié)間長度(倒Ⅰ-倒Ⅳ)均縮短(圖2-F);而且突變體dmc倒Ⅰ節(jié)間的直徑比國麥301要小(圖2-E)。

圖2 突變體dmc和國麥301主要農藝性狀比較
A: 抽穗期突變體dmc(右)與國麥301(左)植株對比,B: 突變體dmc(右)和國麥301(左)穗部對比,C: 灌漿期突變體dmc(右)和國麥301(左)籽粒對比,D: 蠟熟期突變體dmc(右)和國麥301(左)籽粒對比,E: 突變體dmc(下)和國麥301(上)倒1節(jié)間直徑對比,F(xiàn): 突變體dmc(右)和國麥301(左)節(jié)間對比。
?2.2 遺傳背景檢測
利用 SSR標記對突變親本國麥301與突變體dmc的遺傳背景進行檢測比較,結果顯示分布在42條染色體上的431個SSR標記擴增位點均沒有發(fā)現(xiàn)差異性條帶,表明突變親本國麥301與dmc的遺傳背景高度一致。這個結果證明突變體dmc是由國麥301誘變而來的。

M: 分子量標準(DL100);泳道1-3:國麥301,泳道4-6:突變體dmc,擴增產物條帶下是SSR引物名稱
圖3 SSR標記檢測
?2.3 國麥301和突變體dmc的組織學觀察
通過比較不分蘗突變體dmc和野生型對照國麥301的分蘗形態(tài)和顯微結構,發(fā)現(xiàn)dmc中主莖基部的分蘗分化和發(fā)育都受到嚴重抑制(圖4A-E)。在兩葉期,dmc和國麥301在外部形態(tài)上幾乎沒有差異。組織學觀察發(fā)現(xiàn),國麥301的分蘗原基已經(jīng)開始分化(圖4F),但在dmc中未觀察到分蘗原基(圖4A)。在三葉期,國麥301的分蘗芽開始生長,在主莖基部形成兩個一級分蘗(圖4G),而在dmc基部僅形成一個微小的突起(圖4B)。在四葉期,國麥301的主莖變長且增厚,分蘗原基增多(圖4H,圖5A)。同時期的dmc的主莖也伸長,可以觀察到一個微小的分蘗原基在基部(圖4C)。在越冬期,dmc平均株高(在自然狀態(tài)下的最高離地高度)僅為12cm,是國麥301平均株高的一半(圖5B)。組織學觀察發(fā)現(xiàn)在dmc基部只有兩個小分蘗原基(圖4D),但國麥301處于分蘗旺盛期,分蘗數(shù)約為12-14個,分蘗主要由一級分蘗和二級分蘗組成(圖4I)。在起身到拔節(jié)階段,dmc的分蘗幾乎與前期一樣很小(圖4E)。然而,國麥301的分蘗數(shù)已達到最大值(圖4J, 圖5C)。大多數(shù)dmc最終只有一個主莖(圖5C)。
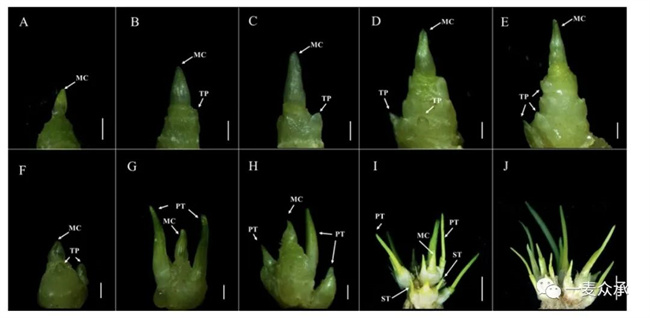
圖4 矮化不分蘗小麥突變體dmc和國麥301的分蘗組織顯微觀察
(A、B、C、D、E:二葉期、三葉期、四葉期、越冬期和起身到拔節(jié)期之間的dmc;F、G、H、I:二葉期、三葉期、四葉期和越冬期的國麥301;J:起身到拔節(jié)期之間的國麥301的部分分蘗芽。MC:主莖;TP:分蘗原基;PT:一級分蘗;ST:二級分蘗。A-H,比例尺=1 mm;I和J,比例尺=1cm。)
?2.4 國麥301和突變體dmc的組織化學觀察
在組織化學水平上,三葉期突變體dmc的分蘗原基與國麥301的分蘗原基有明顯區(qū)別。dmc中通常只有一個小的分蘗突起,被染成淡黃紫色;相反,國麥301通常有兩個分蘗芽,里面包裹著分蘗原基,被染成深黃色。這些表型說明國麥301的分蘗原基蛋白質含量明顯高于dmc。此外,在國麥301的橫切面上,有兩個明顯的黃色區(qū)域,即旺盛分蘗的原基部位。國麥301分蘗原基細胞排列整齊,胞質致密被染成深黃色,呈現(xiàn)典型的分生組織特征。然而,在dmc橫切面中通常看不到分蘗原基及分生組織。總之,dmc分蘗原基少且分蘗原基蛋白質含量低于對照國麥301(如圖4)。
(A:dmc(箭頭)分蘗的縱切面;B:國麥301(箭頭)分蘗的縱切面;C:dmc分蘗基部的橫切面;D:國麥301分蘗基部的橫切面;E:C的放大圖;F:D的放大圖。TP:分蘗原基;TB:分蘗芽。
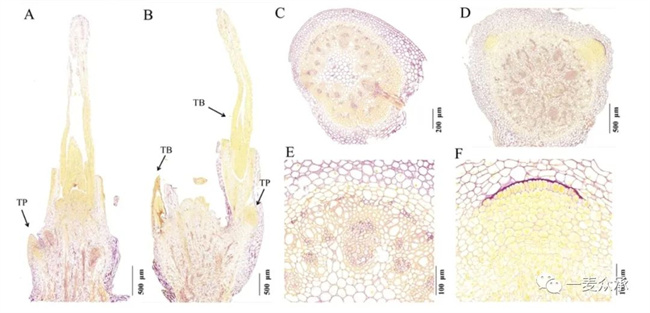
圖5 三葉期的國麥301和dmc小麥分蘗中淀粉(紫色)和蛋白質(黃色)的分布
?2.5 突變體dmc遺傳學初步分析
突變體dmc與中國春進行雜交,對F?進行性狀調查,結果發(fā)現(xiàn)F?分蘗均減少,平均分蘗4.48個,而F?株高介于兩親本之間。初步推測突變體dmc的獨稈性狀可能是由顯性或半顯性基因控制。
表2 突變體dmc、中國春及其F1代的株高和分蘗性

3 小麥提前終止穗分化突變體ptsd1遺傳調控機制研究
?3.1突變體ptsd1表型分析
突變體ptsd1的平均株高和穗長分別為50.75cm和4.19cm,與親本國麥301相比分別降低了21.1%和縮短了60.0%。突變體ptsd1的平均可育小穗數(shù)和單穗籽粒數(shù)為4.50和14.16,與親本國麥301相比分別減少了77.5%和77.9%。千粒重相比于親本降低了19.8%。突變體ptsd1的抽穗期、開花期和成熟期分別推遲了9天、9天和11天。突變體ptsd1主要影響穗長和小穗數(shù)性狀(圖6)。
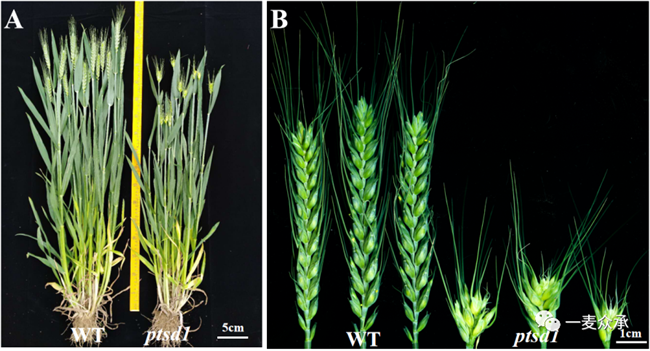
圖6 對照國麥301和突變體ptsd1整株表型(A)和穗部表型(B)
?3.2 突變體ptsd1幼穗分化的超微結構
對照國麥301頂端小穗形成期幼穗超微結構顯示(圖8A),頂端分生組織(im)分化形成最后幾個頂端小穗原基,上部小穗原基已經(jīng)分化形成護穎原基、外稃原基和小花原基凸起,中部小穗中的小花原基已經(jīng)分化形成三個雄蕊原基凸起(stp)和一個雌蕊原基凸起(pip)。突變體ptsd1只有約5個小穗原基正常分化(圖8B),小穗中的小花原基分化形成三個雄蕊原基凸起(stp)和一個雌蕊原基凸起(pip),而上部小穗原基停止分化,最終形成一個個凸起狀結構。經(jīng)過越冬期,頂端分生組織逐漸從營養(yǎng)生長階段向生殖生長階段過渡,頂端圓錐體迅速伸長(圖7 A、A');在二棱期(圖7 B、B'),對照國麥301和突變體ptsd1在都形成小穗脊(spr)結構;在二棱后期(圖7 C、C'),對照的穗軸兩側小穗原基(spp)逐漸開始膨大,首先觀察到中部小穗原基的膨大,然后向頂端和基部延伸,而突變體ptsd1的上部小穗原基停止分化,下部小穗原基逐漸開始膨大;在護穎原基形成期(圖7 D、D'),對照護穎原基(glp)逐漸形成,大約形成12 ~ 14個小穗原基,而突變體ptsd1只形成6 ~ 8個小穗原基;在外稃原基形成期(圖7E、E'),對照在護穎原基上方形成了外稃原基(lemp),大約形成14 ~ 16個小穗原基,而突變體ptsd1只形成6 ~ 8個小穗原基,上部小穗原基仍停留在二棱期;在小花原基形成期(圖7 F、F'),中部小穗的第一個小花原基(flp)分化形成,小花原基位于外稃原基上方,呈半球狀,大約形成16 ~ 18個小穗原基,而突變體ptsd1只形成6 ~ 8個小穗原基,上部小穗原基仍停留在二棱期;在頂端小穗形成期(圖7 G、G'),在這個時期對照除了最后幾個頂端小穗原基分化外,不再形成新的小穗原基,而突變體ptsd1上部小穗原基仍停留在二棱期;在小穗原基分化結束后,對照小花原基逐漸開始分化最終形成雌蕊和雄蕊(圖7H、I、J和K),突變體ptsd1下部小花原基正常分化形成雌蕊和雄蕊,而突變體ptsd1上部未分化的小穗原基沒有分化形成小花原基及隨后的雌雄蕊(圖7 H'、I'、J'和K')。在綠色花藥時期,對照國麥301的上、中和下部小穗中均觀察到正常的雌雄蕊形態(tài)(圖7 L、M和N),而突變體ptsd1上部小穗原基未分化沒有形成雌雄蕊(圖7L'),突變體ptsd1的中部和下部小穗中均觀察到正常的雌雄蕊形態(tài)(圖7M'和N')。觀察記錄穗發(fā)育時期發(fā)現(xiàn),突變體ptsd1頂端圓錐體比囯麥301晚11天進入二棱期,且整個穗分化進程延遲了11天。

圖7 對照國麥301(A-N)和突變體ptsd1(A'-N')的穗分化不同時期形態(tài)
注:A:二棱初期,B:二棱期,C:二棱后期,D:護穎原基形成期,E:外稃原基形成期,F(xiàn):小花原基形成期,G:頂端小穗形成期,H:雌雄蕊原基分化期,I:雌雄蕊原基分化末期,J:外稃完全包裹基部兩個小花,K:綠色花藥時期,L-N:綠色花藥時期對照國麥301的上、中和下部雌雄蕊形態(tài)。A'-N': 突變體ptsd1與對照發(fā)育時期一一對應。spr,小穗脊;spp,小穗原基;spd,小穗分生組織的圓頂;glp,護穎原基;lemp,外稃原基;flp,小花原基;ts,頂部小穗;an,花藥。紅色括號標注的是從二棱期開始突變體ptsd1上部小穗原基不分化。小麥穗發(fā)育時期參考文獻。

注:Im,花序分生組織;stp,雄蕊原基;pip,雌蕊原基
圖8 對照國麥301(A)和突變體ptsd1(B)頂端小穗形成期幼穗超微結構
?3.3 遺傳分析
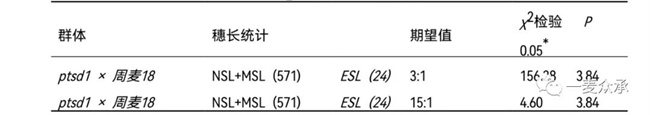
突變體ptsd1與周麥18雜交組合,正反交F?穗長正常(圖9A-D),表明控制穗長性狀位點為隱性基因。“ptsd1 × 周麥18”雜交F?,穗長性狀分離為三種表型,正常穗(NSL)、中等縮短穗(MSL)和極端縮短穗(ESL)。其中,正常穗的長度與親本周麥18穗長相當,中等縮短穗的長度介于周麥18和突變體ptsd1之間,極端縮短穗的長度與突變體ptsd1相當。統(tǒng)計595個F?單株穗長表型發(fā)現(xiàn),正常穗、中等縮短穗和極端縮短穗分別為503、68和24個單株(表3)。假設穗長由一對隱性基因控制,統(tǒng)計正常穗和中等縮短穗(NSL和MSL)為571株,極端縮短穗(ESL)為92株,χ2檢驗結果顯示χ2 > P0.05,不符合孟德爾單基因遺傳3:1的分離比,因此突變體ptsd1不是單基因隱性遺傳。假設穗長由兩對隱性基因控制,χ2檢驗結果顯示χ2 > P0.05,不符合15:1的分離比,因此突變體ptsd1也不符合孟德爾兩對獨立的隱性基因遺傳。
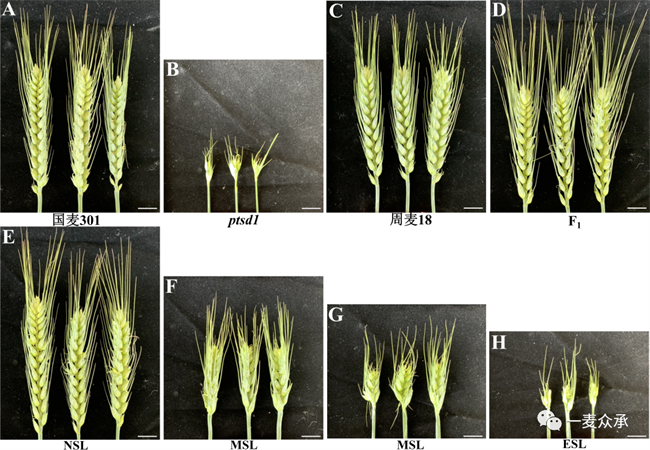
圖9 對照國麥301(A)、突變體ptsd1(B)、周麥18(C)、ptsd1與周麥18正反交F?(D)以及后代F?或F2:3穗部形態(tài)(E-H)。
表3 F?群體穗長性狀遺傳分析
?3.4 結論
周麥18和中國春分別與突變體ptsd1雜交,F(xiàn)?穗型都正常,表明ptsd1控制穗長性狀是由隱性基因控制的。在F?和F2∶3群體中,正常和突變植株的分離比不符合經(jīng)典孟德爾遺傳學的1對隱性基因或2對獨立分配、自由重組的隱性基因遺傳的理論分離比。表明突變體ptsd1中,穗長性狀不是由一對隱性基因也不是由兩對獨立遺傳隱性基因控制。660K芯片SNP標記連鎖分析和SSR標記連鎖分析表明,在5A染色體的短臂和長臂上各有一個突變位點。因為這兩個突變位點在同一條染色體上,存在基因連鎖,所以遺傳分離比不符合15:1。為此,ptsd1是首次發(fā)現(xiàn)的在5A染色體上同時存在兩個基因控制穗分化的突變體。未來,我們將把這兩個基因分解到不同的作圖群體中,從而對每個基因進行圖位克隆。
基于目前的研究結果,構建了突變體ptsd1的遺傳模型(圖10)。將兩對基因的四種基因型分別用A、a、B和b代替,如A:Ptsd1-5AS,a:ptsd1-5AS,B:Ptsd1-5AL和b:ptsd1-5AL。突變體ptsd1攜帶兩對隱性基因aabb,正常穗長植株攜帶兩對顯性基因AABB,雜交F?基因型為AaBb。F?自交兩代,在F2∶3篩選穗長分離的家系。根據(jù)F2∶4的分離情況,確定F2∶3家系單株是否純合。篩選純和的正常穗基因型AABB和極端縮短穗基因型aabb純合株系構建BSA混池,本研究篩選獲得的連鎖標記都是基于這兩個混池。
將兩個基因分解到兩個遺傳群體的方法有以下兩種:
1)篩選穗長性狀分離的雜合F2∶3植株,根據(jù)連鎖標記鑒定F2∶3中基因型為aaBb和Aabb的單株,自交一代得到F2∶4家系群體為一個基因純合,另一個基因分離的遺傳群體;
2)篩選穗長性狀不分離的純合F2∶3植株,根據(jù)連鎖標記鑒定F2∶3中基因型為aaBB和AAbb的單株,與突變體ptsd1回交一代得到基因型為aaBb和Aabb的單株,自交后獲得單個基因的遺傳群體。

圖10 突變體ptsd1的遺傳模型
來源:一麥眾承





![[人民日報] 我國篩選出耐鹽堿種質資源1100多份](http://m.gowu797.com/file/upload/202411/08/092303811.jpg)

